
Note to readers: Biological Expositions is a series of blog-posts each of which is equivalent in content to a book chapter. If a bio-bullet is likened to a starter, our routine blog-post could be seen as a light lunch and a Biological Exposition blog-post as a three course meal. We look forward to your comments on this series.
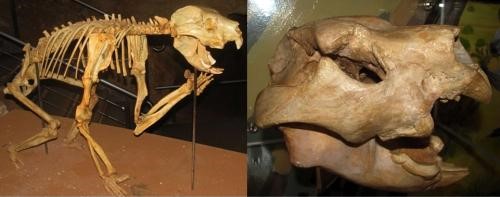
The so-called marsupial lion (see Figure 1) is the largest carnivorous marsupial known from the fossil record: a giant relative of koalas that presumably ate giant relatives of wombats and kangaroos in prehistoric Australia. This metatherian or ‘non-placental mammal’, celebrated as the Antipodean answer to the sabre-tooth felids of other continents, became extinct about 40 thousand years ago, when its large prey died out and the human species became the main predator in Australia. Although all carnivorous mammals on land have carnassial cheek-teeth capable of snipping flesh when butchering a carcase, those of the marsupial lion were particularly impressive, being much larger, simpler and more prominent in the mouth than the carnassials of felids, canids, or bears. Indeed, the sheer length of its carnassial blades exceeded even that of the polar bear, an animal that weighs three-fold more than the marsupial lion did and eats whales by dissecting massive pieces of hide and blubber. But the marsupial lion differs from all other carnivorous mammals on land in lacking prominent canine teeth. Instead of explaining this anomaly, scientists studying the fossils of this species have made a case of simple substitution – claiming that not only were the prominent but blunt incisors of this species somehow sharp enough to puncture hide, but that such puncturing was deep enough to kill the prey.
In this essay we strip the prevailing view of the marsupial lion down to the bare bones of its questionable assumptions. Although we acknowledge that this largest of terrestrial predators in prehistoric Australia may have – in the largest individuals – reached the average body mass of the female lion, we suggest that the ‘lion’ in marsupial lion is a misnomer. Indeed, we will argue that it was less like a felid than like a giant, modified browser which had evolved to eat living flesh instead of eucalyptus leaves.

From our viewpoint at the Bio-edge, the mistake made by palaeontologists in the case of the marsupial lion is based on a pseudo-scientific credo that has become too common in biology: namely that the evolutionary process is partly or largely accidental and that, given the limited genetic plasticity of organisms, natural selection can only tinker with what is available in a given lineage rather than invent whole features. Those who would invoke accidental explanations for the anomalous dentition of the marsupial lion appear to have overlooked a wider principle. This is that the primary process in filling any ecological niche on a continent is the recruitment of the most suitable applicant – the one having what amounts to the best prior qualifications and experience within what is usually a diverse fauna of candidates. Following this recruitment of a suitable lineage of organisms, a secondary process is the detailed modification of this ancestor by further genetic mutation and selection. We realise that the Darwinian focus tends to be on this subsequent stage, i.e. the one in which natural selection fine-tunes an incumbent into fitness for the precise niche in question. However, it is sometimes overlooked that this, as it were, ‘on-the-job training and practice’ depends on an initial selection from a wide faunal field of job-applicants. And, even if the best of a bad bunch of available recruits is far from perfect, the remarkable examples of evolutionary convergence in biology text books remind us that genetic plasticity can still shape – by natural selection – such flawed recruits into eventual fitness for the niche in question.
So join us as we take a virtual tour in an imaginary Museum of Prehistoric Australia, to reveal the evidence and insights that have led us to the view that the marsupial lion may need a new identity. At Display one, we face up to a perplexing contrast: the apparent failure of our Australian metatherian, the marsupial lion, to develop large canines, compared with the success of a South American metatherian, the pouch-sabre – possibly the epitome of the evolution of large canines. At Display two, we turn the spotlight on the scientific and cultural biases that have previously dogged the quest for the true nature of the marsupial lion and the real functions of its peculiar teeth. Display three introduces the idea that a combination of ‘pouch-robbing’ and ‘cookie-cutting’ may have been a strategy appropriate for the marsupial lion, based on insights into how the slow growth of marsupial prey might have constrained the niche available to the largest predatory mammal in Australia. Proceeding to Display four we scrutinise crucial similarities between this mammal and the living koala. Display five reveals that even the historical dingo was sustained by what was effectively ‘pouch-robbing’ of its prey-base of modern kangaroos. And our final display draws the evidence together in a new light as we travel back in time to 50 thousand years ago, to visit the real marsupial lion in its real context.
So, let us open our imaginary tour of this palaeontological riddle at Display one. Here we can scrutinise the accepted interpretation of the marsupial lion: a dashing slaughterer of diprotodons and other marsupial megaherbivores, equivalent to the modern lion or even to extinct sabre-tooth felids. One does not have to be a biologist to spot the obvious shortcoming of the marsupial lion in living up to this reputation: even a glance at the skull will show that it lacks any teeth capable of stabbing prey. This is because its canines are so small and blunt that they are easily overlooked (see Figure 2), and although several other teeth in its jaws are prominent, none is a separate, puncturing tooth, and no combination of teeth could possibly function as a canine typically does. In particular, the left and right lower incisors grow side-by-side so closely that they do not present separate points; and their united apex is so rounded that the net effect is that of a blunt instrument which is unlikely to have been effective in piercing the hide of large animals. Whereas we doubt that the blunt incisors and small canines were able to kill large prey, enlarged bones on the first digit do indicate the likelihood of a talon-like claw on each forepaw – perhaps acting as a substitute for the missing canine. However, even this claw seems uniquely configured among large carnivorous mammals, living and extinct, in being extremely differentiated between the first fore-digit and all the other digits; the question of its real roles in conjunction with the unique dentition of the marsupial lion remains unanswered.

Further doubt is cast on the leonine image of our marsupial by several contrasting skulls in the same display case, starting with the pouch-sabre of South America (see Figures 2 and 3). Belonging to a lineage of mammals just as primitive as the marsupial lion, this species shows the potential of metatherians to evolve canines almost to a fault – as the upper canines of the pouch-sabre were longer than those of most sabre-tooth felids known from the global fossil record and also transcended them in growing and self-sharpening beyond adulthood. Indeed, the pouch-sabre arguably presents the ultimate example of canine development at the expense of the rest of the dentition. In contrast to the marsupial lion, it lacked incisors altogether and even the shearing premolars expected in meat-eating mammals were remarkably reduced. In this same display, skulls of additional metatherians show that modest but perfectly respectable canines were already present in a carnivorous lineage of Australian marsupials which survives to the present day – the dunnart order – which begs the question of why a more truly leonine marsupial did not evolve from this lineage instead of the dentally anomalous relative of the koala that is at the centre of our quest for the truth.

Our opening display has shown that evolutionary recruits, possessing suitable canine teeth, were in fact available in Australia all along. It seems that natural selection did not, after all, need to reinvent; it could have tinkered and still produced a version of a marsupial lion with long, puncturing canines had there been a niche for such an animal. As we continue our virtual tour, our scepticism grows: could it be that the largest carnivorous metatherians in Australia and South America diverged not by accident, but instead because Australia presented a unique set of environmental challenges for which typical canines were inappropriate?
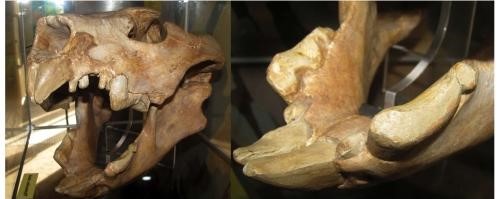
We turn now to Display two, which uncovers the subconscious motives of the scientists who have interpreted the fossil record. Instead of directly confronting its paradoxical dentition, palaeontologists have for one-and-a-half centuries presented obtuse – and seemingly biased – interpretations of the marsupial lion. They have described its blunt upper incisors as caniniform’ and their illustrations have misleadingly suggested that the incisors, both upper and lower, are acute enough to resemble canines – either because the skull is photographed in full profile (see Figure 4) or because reconstructions of the living animal exaggerate the separateness and sharpness of the incisors. But any frontal view of the real skull (see Figure 5) will show that left and right incisors are bluntly united, much as they are in the herbivorous members of the same diprotodont order, including koalas. Given that the gap between left and right incisors is clearly insufficient to allow stabbing, how objective has the assumption of a lion-like use of the teeth really been? And to what degree has the acceptance of this assumption by the educated public been swayed by the emotive power of large predators?
In this case, evolutionary theory may have fallen prey to a pseudo-scientific double standard. On the one hand, palaeontologists seem to accept as a case of evolutionary convergence the similarity in dentition between the thylacine of Australia – the largest of the dunnart order and the second-largest meat-eating marsupial ever known – and the fox of the Old World. And indeed this would seem reasonable, particularly because, of all the body parts of mammals, it is likely to be the teeth – the cutting edge of survival – that are least constrained by genetic impediments to evolution. However, on the other hand, the same palaeontologists imply that evolutionary plasticity has been insufficient for natural selection to enlarge the small canines found in the ancestor of the marsupial lion by mere tinkering, and instead prefer to imagine this dentition emulating the quite different dentition of the felids that occur widely beyond Australia. Surely one tenet negates the other: if a niche called for the evolution of canines rivalling those of the lion, then the availability of canine-bearing carnivorous marsupials would have predisposed the dunnart order, rather than the diprotodont order – to which koalas belong – as the recruit for further evolution?
Is the subconscious drama of predation, then, so powerful that even the best scientists have lapses of objectivity? And is the current celebration of the marsupial lion partly a reflection of a misdirected Australian jingoism that subconsciously echoes the line ‘You call that a knife?’ from ‘Crocodile Dundee’?
With our doubts growing along these lines, we now move to Display three. This shows us how ‘pouch-robbing’ and ‘cookie-cutting’, rather than stabbing adults to death, may have been appropriate tactics for an animal with the dentition of the marsupial lion in the Australian context.
A clue to the puzzle of the real ecological niche of this largest of meat-eating marsupials is the observation that marsupials generally have slower metabolism and growth than expected for comparable eutherian mammals. Assuming that the erratic climates and poor soils of modern Australia applied also in the prehistoric period, it would make sense that the giant wombat and other large herbivorous marsupials, which became extinct around the same time as the marsupial lion, were slow-growing compared to eutherians of similar body mass found on other continents. This clue points to a crucial principle in predation: that slow growth of prey will necessarily constrain the rate of consumption of any top carnivores dependent on such a prey-base. Indeed, the trickling productivity of herbivores in prehistoric Australia is unlikely to have matched the appetites of large felids, which – despite their apparent laziness – have rapid metabolism and growth and require large amounts of rich food. It is perhaps unsurprising then that the fauna of Australia did not contain real felids or even truly felid-like marsupials. Instead, the real carnivorous niche offered by the ungenerous and unreliable Australian environment might have been that of ‘browsing’ from living animals while killing adults as seldom as possible in the interests of sparing the breeding stock of the prey.
The principles presented in Display three suggest that, if prey can somehow be ‘browsed’, the slow growth of herbivores need not be an insurmountable problem for top carnivores. One browsing option might have been, for example, the harvesting of joeys rather than adults, thereby conserving the prey-base. Along similar lines, a mammalian predator might have attempted in some way to emulate the cookie-cutter shark, which contents itself to bite chunks of skin and flesh out of tunas and dolphins, leaving their bodies to heal. This shark, although a parasite as much as a top carnivore, hints at the plausibility of a strategy that spares the breeding adults of a prey species to provide another offspring, or another bite, on a later occasion.
Extracting offspring by means of a combination of claws and incisors might have allowed the marsupial lion to rob the pouch while sparing the mother. The supposed talon on each forepaw and the pincer- or plier-like paired incisors could theoretically have been used to force open the maternal pouch and to prise out the joey. In addition, the distinctive combination of digits and dentition of the marsupial lion could have been used to secure flaps of skin and muscle for quick shearing from the rump, and to snip other pieces from adults of the largest herbivorous marsupials – such as sections of the fleshy tails of giant, slow kangaroos.
As we continue to ponder Display three, it occurs to us that, although no carnivorous mammal living today could be called a ‘cookie-cutter’, hints in this direction are evident. For example, hyenas amputate the tails and ears of various large animals while chasing their prey or by sneaking up to resting animals at night. Such cropping fits in with the fact that the tail, although used to store fat in certain species, is one of the most expendable parts of the mammalian body and is routinely severed in the domestic husbandry of sheep and dogs. And we also note the possible significance of the fact that hyenas do not have particularly well-developed versions of the bone-crushing molars found in the wolf and wolverine – rather relying on large, specialised premolars that are blunt enough to crush large bones but sharp enough and close enough to the front of the mouth to function as amputators.
Based on living species such as hyenas, then, it does seem possible that the marsupial lion could have ‘browsed’ herbivores without killing or excessively crippling them. The unusual combination of extremely prominent carnassials and a lack of bone-crushing molars – together with the small, obscure canines – is certainly consistent with an unorthodox method of biting. Although its incisor and carnassial rows were not finely serrated enough to emulate the trimming of small pieces of flesh from bone that is so typical of felids, the superlative carnassials of the marsupial lion (see Figure 5) can be interpreted as specialised for the vivisection of sizeable body parts before the large victim could defend itself. And a consistent explanation of the subordinate nature of the canines in our paradoxical marsupial would be that prominent canines would be redundant – or even obstructive – in the hasty act of theft and mutilation that we envisage. By contrast, the large claws and sturdy pincer- or plier-like paired incisors would help by securing a body part immediately prior to any shearing action.
Pondering the true nature of the marsupial lion, it seems likely that its appetite would have been modest if – as expected – it had the slow metabolism and reproduction typical of related marsupials. Another reasonable inference, based on the sparsity of fossils of this species, is that it was nowhere abundant, and solitary rather than gregarious. A lone individual might not have been able to consume the carcase of even a half-tonne diprotodon before this rotted or was scavenged. On this basis we suspect that the marsupial lion would have wasted food by the act of killing adult herbivores, making browsing of herbivores a more prudent strategy than killing them.
The realisations we have made in Display three also support the idea that both the marsupial lion and its prey species grew slowly compared to the eutherian prey species found in the Old World. Indeed, a pattern of slow growth of herbivores has been consistent across Australia because marsupials consume resources more slowly than most eutherians do – as explained by characteristically erratic climates and poor soils rather than simply an accident of continental isolation. Even the modern kangaroos and wombats, which have been resilient enough to survive the arrival of the dingo from Asia, replace individuals infrequently in the long term compared to ruminants of similar body mass. The average kangaroo, for example, consumes energy at only a quarter of the rate of the average sheep. From the point of view of a predator, this translates into only a quarter of the flesh available for harvesting in any given period.
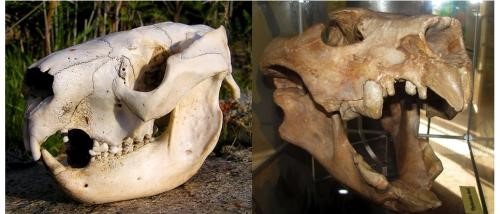
Moving on to Display four, we are grounded once again in the observed reality of the modern fauna. Palaeontologists seem reluctant to acknowledge that the closest living relative of the marsupial lion appears to be the koala, that quintessential Australian herbivore with its specialisation on eucalypt leaves and a brain too diminutive even to fill its small cranium. It has, however, been established that the cranial capacity of the marsupial lion was only 40% of those of felids of similar body mass, indicating that it retained the modest cognitive ability of its koala lineage, as well as a similar configuration of limb proportions. Although the koala and marsupial lion share a similar arrangement of incisors and canines (see Figure 6), it is remarkable that this comparison has not informed the puzzle of the dentition of the marsupial lion and that nobody seems yet to have studied precisely how even the familiar koala uses its own front teeth. We have observed that the koala hardly seems to need the incisors for browsing because it clips the petioles of eucalypt leaves, one by one, using its premolars directly. What it can do with its incisors is fight and defend itself, but even in those capacities the incisors seldom draw blood and function as pincers or pliers rather than cutters or stabbers. Did the marsupial lion perhaps use them in a similar fashion?
The affinities between the marsupial lion and the koala also suggest that the guts of the marsupial lion were possibly odd among carnivores, including the thylacine. This is because the koala has bulky intestinal chambers for fermentation of plant fibre. The inference that the marsupial lion also had bulky intestines is consistent with a new, adaptive view of the marsupial lion, because if it really was an ‘animal-browser’ then much of its diet might have been a different type of fibre: the keratin that makes fur and reinforces hide. We will never know the exact nature of the gut of the marsupial lion, but its phylogenetic affinities with koalas suggest that it might be fibre-digesting. Although no living vertebrate is known to use a microbial symbiosis for the digestion of keratin as a source of energy, such a possibility is suggested by the fact that many types of bacteria have been found to produce the unusual enzymes required in the recycling of keratinous wastes from livestock farms.
Moving to Display five in our virtual museum, we briefly observe the dingo, which itself turns out to have been something of a pouch-robber during the historic period. This canine subspecies was able to cope with a relatively unproductive prey-base of marsupials because it was small enough to meet its needs from a continual trickle of juvenile kangaroos produced from its attempts to hunt down adult mothers. Meals from joeys remained just large and frequent enough to allow this wild canine a staple diet of animal matter – at least in conjunction with scraps from Aboriginal camps. Indeed, there is a significant anomaly of marsupial behaviour which evidently helps the dingo inadvertently to harvest the populations of kangaroos sustainably rather than deplete them; the anomaly being that when chased to a point of distress – but not necessarily injury – the mother kangaroo relaxes the muscles of the pouch, allowing the juvenile to tumble out and be sacrificed . Even if the joey escapes predation, it is subsequently rejected by the mother and thus doomed anyway. Such jettisoning may make sense in terms of lifelong reproductive effort when one considers that the dingo needs to be appeased to the point where it does not hunt down and kill adults at an unsustainable rate. Presumably the marsupial lion lived by the rule evident in the predation of kangaroos in historical Australia: namely that any consumer of slow-growing herbivores needs to harvest judiciously rather than wantonly to make ends meet.
At the end of our museum tour, we have an opportunity to view prehistory with new eyes, our marsupial lion portrayed in a plausible alternative reality. Our final display is a diorama of a scene 50 thousand years ago in semi-arid Australia, showing a diprotodon dozing under a tree and a marsupial lion looking on with the slit pupils and somnolence of a koala but without the same innocence. In this setting our newly informed imaginations can animate the scene. We see that the locomotion of the marsupial lion is plantigrade and slow, and that it attempts neither to run down prey nor to scavenge bones as hyenas do on other continents. Instead, we can see it creeping up on marsupial victims and pouncing to take a mouthful of hide and flesh before retreating; we can see it approaching sleeping members of large, slow-moving species closely enough to extract offspring from their bulging pouches by means of its incisors and claws; and we can see it harassing evasive mothers until they jettison their vulnerable joeys. The inferiority of its canines and the bluntness of its projecting incisors suggest that the marsupial lion, far from attempting to stab prey with any of its teeth, retained the approach of using its premolars to clip off food items but switched from browsing plants to, as it were, browsing animals. And even the snarl invariably painted into artistic reconstructions of the species is absent, its lips pursed until the moment of violation, for there is no reason to display the teeth.
As we leave our Museum of Prehistoric Australia, we are mindful that the pie of ecological niches has been sliced in unusual ways in Australia, perhaps in response to a unique combination of climate, soils, and fire regimes. Here we have the largest mammals that hop (kangaroos), the largest herbivores that dig complex tunnels (wombats), the only specialised nectar- and pollen-eating mammal that neither flies nor glides (honey possum), and the only specialised ant- and termite-eating mammal that lays eggs (short-beaked echidna). Given that all these forms share a consistent trait of slow growth relative to their closest eutherian counterparts on other continents, it is possible that Australia can also claim the only specialised ‘browsing’ meat-eater among mammals. The usual assumption that the aberrant teeth and claws of the marsupial lion merely emulate the typical teeth and claws of large felids seems not only to miss the significance of evolutionary recruitment from a herbivorous lineage rather than the carnivorous lineage available among the ancient marsupials, but is also incongruent with the unique challenges and opportunities presented by the Australian environment. If there was an odd carnivorous niche to fill, it may be no accident that an odd ancestor was recruited to fill this niche, and that there was evidently no selective pressure to enlarge the small canines retained by the incumbent.
Although the term ‘natural selection’ is often taken for granted, it turns out to have two meanings. In the recruitment process, the selection is of an ancestor that comes with all the biological advantages conferred on it by previous evolution; in the subsequent modification of detail, the selection is instead among individuals based on genetic mutation. A ‘meat-browsing’ strategy would fit the fact that the marsupial lion was not derived from the dunnart order, with its pre-adapted canines, but instead from the same herbivorous lineage as its prey in the diprotodont order. This derivation suggests that, despite a switch in diet from vegetable to animal, the use of the front teeth remained consistent with a browsing approach to carnivory. There are precedents for such a shift by means of recruitment, because other switches by herbivorous lineages to carnivory, and vice-versa, are known to have occurred here and elsewhere. An example is the largest native species of rodent in Australasia, the rakali, which belongs to the same lineage as a specialised herbivorous rodent in Australia. The rakali is amphibious and eats birds, large invertebrates, and other animal matter to the virtual exclusion of plant matter, despite the prevalent use of sharp incisors in other rodents for gnawing plants. And as a familiar example of the converse switch, from carnivore to herbivore, we have the evolution of the specialised, bamboo-eating giant panda from ancestors in the carnivore order.
Although our interpretation of the marsupial lion as a pouch-robber and cookie-cutter is unconventional, it hinges on the fact that Australia is not just the most isolated continent – alternatively the largest island of all – but is also characterised by droughts generated by temperature fluctuations in the Pacific Ocean as well as widespread and long-standing poverty of phosphorus and other nutrients owing to geological senility. One cannot understand Australia without appreciating that, as much as there are constraints in its faunal origins, these climatic and edaphic hardships and the consequent poor productivity have also shaped its flora and fauna. Our rationale is that this marginal Antipodean environment resulted in the evolution of one of the most economical of all mammalian carnivores – partly by virtue of the recruitment of a lineage already adapted to the fibrousness and nutrient-poverty of Australian plants as food.
History has a subtle way of repeating itself. The European settlers in Tasmania failed to conserve the thylacine partly because they tended to view it as a default canine, little more than a primitive – and possibly inferior – version of the wolf. And, just as this mindset led to the extermination of the largest meat-eating marsupial surviving to modern times, it may also have eclipsed the real nature, known only from its bones, of the largest meat-eating marsupial of all time. This would not be the first time in science and life that false assumptions have outweighed the real evidence. Palaeontologists and the public have ostensibly fallen prey to an assumption that a niche for large felids must have existed in Australia as on other continents, and consequently that the marsupial lion was the most similar metatherian that a genetically limited system could produce. We trust that our constructive scepticism will expand the image of the marsupial lion, rather than diminish it. It is understandable, given the special capacity of felids to combine human appeal with bestial ferocity, that Australians, educated in the traditions of the Old World, have harboured subconscious misgivings about living on the only inhabited continent that lacks any big cat. However, by claiming against the evidence that Australia was, in the past, home to an iconic predator, the prevailing view has ironically missed something special about the Australian bestiary. For the land Down Under is distinctive not only in its accidents of history but also in the environmental regime which – according to any solid evolutionary theory of cause and effect – is the ultimate adaptor in a fauna. All continents offered pre-existing canine teeth for the niche of top carnivore. But, in the case of Australia, it appears that puncturing teeth were rejected as an error by the trial of evolution.
Links:
See misleading portrayal of sharp, canine-like incisors in painting by Martin Thompson here.
See the video clip of Crocodile Dundee’s classic line here.
See the video clip of a koala in action here.
Acknowledgments:
Our sincere thanks to Roman Uchytel (www.prehistoric-fauna.com) for permission to present his world-class reconstructions in this Exposition, including the image of the marsupial lion (Thylacoleo carnifex) on the title page.