
Note to readers: Biological Expositions is a series of blog-posts each of which is equivalent in content to a book chapter. If a bio-bullet is likened to a starter, our routine blog-post could be seen as a light lunch and a Biological Exposition blog-post as a three course meal. We look forward to your comments on this series.
A peculiarity in the body plan of fishes seems to have been widely under-looked. This is that the pelvis of certain fishes has been brought up from where it belongs, near the vent and the start of the tail, to a position ‘above the shoulders’ in human anatomical terms. Imagine carrying your pelvic girdle on top of your shoulders and having your legs dangling above your arms. This is the sort of perversity that can go unnoticed in fishes.
Two classes of animals are actually confused under the vague term ‘fishes’: these are the bony fishes and the cartilaginous fishes, which are as different from each other as birds are from mammals. We refer in this blog-post particularly to the bony fishes, a group containing almost as many species as all tetrapods (mammals, birds, reptiles, and amphibians) combined. This is our focus because the cartilaginous fishes, consisting of chimaeras, sharks, rays, and skates, differ little in their pelvic placement relative to mammals, reptiles, and amphibians, whereas the bony fishes have pelvic perversion to various degrees, and in some respects show a convergence with birds.
To those readers who see fish anatomy as a rather cold topic: the pelvis is the scaffold of our loins and our hips indicate our gender. It is through the pelvic girdle that we are ecstatically conceived and excruciatingly born. Indeed, the pelvis would seem to rival the skull in embodying what matters most to vertebrates.
Fishes, however sexually streamlined, also have a definite pelvic dimension, with the South American lungfish being an extreme example. Males of this particular fish have threadlike pectoral fins and females have threadlike pelvic fins – neither of which is particularly remarkable. But the thought-provoking thread is that the pelvic fins of the male develop during the breeding season into a pair of feathery appendages. These fins have no apparent locomotory function, but release oxygen from the blood of the fish into the water and are apparently used to oxygenate the eggs in the nest. In this way, the father conveys some of the oxygen he normally gulps at the surface to his offspring via a pelvic structure, an odd involvement of the hips in paternal care.
In order to appreciate the distortion in the appendicular skeleton that is taken for normal in bony fishes, you need to recognise two anatomical points of reference: the pectoral girdle and the vent. Once these are established, a quick circuit of any aquarium – even distracted by having the children in tow – can expose the pelvic vagaries of a range of fishes.
The pectoral girdle of bony and cartilaginous fishes corresponds to the human shoulders, and can be found roughly where expected: between the head and the viscera. Its location is perhaps one of the most consistent features of fish anatomy, even though the pectoral fins show extreme variation in size and are used variously for propulsion (seahorses), gliding flight (flying fishes), camouflage (stonefishes), flashing warning colours (gurnards) and even probing the sea floor (the anterior pectoral rays in gurnards), as well as braking and steering in most fishes. Where the consistency fails is in certain eels, such as morays and swamp-eels, which have lost their pectoral fins during the evolution of the most serpentine body form attained by fishes.
The vent, which includes the anus but is actually a cloacal opening, is located in most bony fishes at the junction of the ‘body’ and the ‘tail’. This orifice is shared by the faeces, urine, and reproductive products and its position is not as consistent as that of the pectoral girdle. Although the normal position of the vent suffices as a reference point for our purposes, possible confusion between ‘vent’ and ‘ventral’ warrants an explanatory footnote.
There is an easily overlooked incongruity in the word roots for ‘vent’ and ‘ventral’: in many mammals, ‘ventral’ does not correspond with the location of the ‘vent’. For example, picture a dog, cat, or cow: its vent is its anus, located at the top back corner of the body. But its ventral surface is its belly, quite separate from its anus.
Given the normal position of the vent in fishes, if you draw a line from this orifice to the throat you trace the axis along which the pelvic girdle has shifted various distances in various lineages of bony fishes (see the cover illustration of this blog-post). Anyone who has prepared a fish for filleting will recognise this as the dotted line for that gut-wrenching knife-slash.
Now move along this line in your mind’s eye and try to locate where you think the pelvic girdle should lie. You would probably be wrong in your guess, because relatively few of the approximately 28 thousand species of bony fishes have the pelvis where it might be expected based on cartilaginous fishes, amphibians, reptiles, and mammals. Indeed, in most bony fishes, the pelvis has been displaced forward – in human terms to our waist and beyond.
Pelvic shift one occurs in sturgeons (see Figure 1) and some catfishes. Although this is the minimal shift within bony fishes, the pelvic fins are farther from the vent than in any cartilaginous fish.
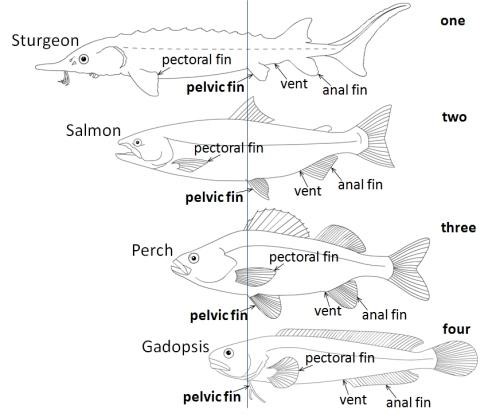
Pelvic shift two occurs in salmons, flying fishes, and herrings. A considerable distance is maintained between the pelvic and the pectoral girdles, but the pelvic fins have shifted to an abdominal position. For perspective, imagine our private parts where they are, near the end of our spine, but our pelvis and its attached legs having slid up to our waist. Pelvic shift three occurs in most species of bony fishes, in which the pelvic fins are located right next to the pectoral fins. In human terms, this would mean having our pelvis either snugly under our armpits, or actually replacing our collarbones. This arrangement has become normal for the wonderfully diverse and colourful fishes that flutter about on coral reefs, such as angelfishes, damselfishes, and wrasses, as well as the predatory trevallies and their relatives which dash about like quicksilver in the adjacent waters.
In pelvic shifts one to three, the pelvic fins tend to retain a consistent shape and size despite the progressive shifts. However, once the pelvic girdle actually passes the pectoral girdle, the pelvic fins are first modified and then lost even though the internal pelvic bones may be retained.
In pelvic shift four, the pelvic fins have moved ahead of the pectoral fins to be tucked up just behind the throat. This occurs in several groups of bony fishes, some long-bodied and others squat, including gadopsis, otherwise known as the Australian river blackfishes (see Figure 1), cods and their relatives, blennies, anglerfishes, oarfish, gobies, and mudskippers. Referring back to our human form, imagine that the hips of all these fishes have climbed past the waist, armpits, and shoulders to the throat, being worn like a necktie or taking the place of our larynx (Adam’s apple).
Please reflect for a moment that fishes which fall under pelvic shift four often appear quite normal to the human eye. Cods and their relatives, for example, are not viewed as contorted freaks but rather as appetising, standard grey fishes. However, their pelvic and pectoral bones have actually connected. The girdles are not just close to each other, but are partly fused, as a human pubis might fuse with a human collar bone. Similarly, in the oarfish, the longest of all bony fishes with a maximum length up to 17 metres, the pelvis is located near the gills, yet most human observers would not remark on the pelvic oddity of what is probably the longest distance between any pelvis and any tail tip on Earth, including the blue whale. The peculiarity of the pelvic fins of the oarfish goes further because they are so long and narrow that they seem dysfunctional. By contrast, some gobies and mudskippers seem to place extreme reliance on their pelvis because the left and right pelvic fins are fused into a single sucker, allowing the fishes to cling to a substrate or to locomote above water level at low tide. And to add a finishing flourish to pelvic shift four, most species of the bottom-dwelling, sluggish anglerfishes do not leave the water but use their pectoral and pelvic fins virtually as limbs. In fact, anglerfishes move in the slow-motion equivalent of a cantering gait, in which the pectoral members are swung forward in unison while pivoting on the pelvic members.
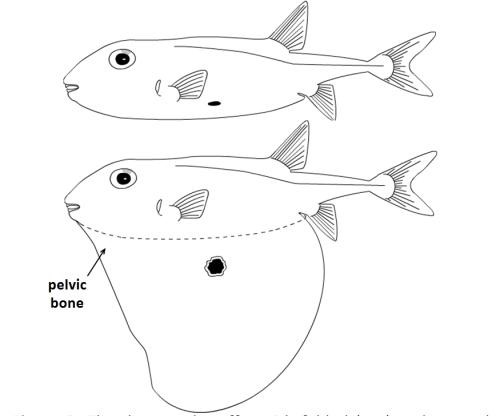
In pelvic shift five, the pelvic bones, which remain well-developed despite the loss of the pelvic fins, have moved to the throat or even in front of it. One example is the leatherjackets, in which instead of pelvic fins there are mere spines. Paradoxically, it is with this extreme pelvic shift that the internal pelvic bones have become so mobile and muscular in certain fishes that they can change the ventral geometry of the body. When erect and locked, the pelvic mechanism gives refuge to leatherjackets by jamming them in secure nooks among rocks. The ultimate erection of this kind occurs in the threetooth puffer, which only partly inflates the body with water and, unlike other pufferfishes, also has a pole-like internal pelvis that rotates downward to support an extended but not inflated flap (see Figure 2). Here is what looks like the fish equivalent of a large dewlap hinged at the throat – but is actually a pelvis, shifted to beneath the head and containing what must be one of the largest pelvic bones in any fish, proportional to body size.
We do not know enough about the internal pelvic bones to place certain groups of bony fishes – which have lost the pelvic fins altogether – along the series of pelvic shifts we have described. Examples are seahorses, eels, swordfish, hairtail, toadfishes, rock ling, and the ocean sunfishes and pufferfishes. Some of these may even have lost the internal pelvic bones, and studying the peculiar modes of locomotion of all of them would keep ichthyologists busy for decades.
Based on this fresh look at familiar silhouettes, bony fishes may tend to resemble birds in the arrangement of their fins and limbs. In both birds and bony fishes, gravity is somewhat alleviated by internal air sacs or a buoyant medium. This allows some loss of bodily support by the axillary skeleton and, in the case of fishes, dislocation of the pectoral and pelvic girdles from the spinal column. There may thus be overlooked homologies and analogies between bony fishes and birds in the arrangements of fins and limbs.
Most birds have a short body between the forelimbs and the hindlimbs, the legs being arranged just below the wings in normal standing or perching positions. This means that the limbs of birds tend to spring from the centre of gravity of the body, an arrangement possibly better suited to acceleration and deceleration than is the separation of limbs typical of lizards, crocodiles, or mammals. The shift of the hindfins of bony fishes towards the forefins similarly serves to locate the steering and braking ‘limbs’ close to the centre of inertia, separating bony fishes from cartilaginous fishes. In bony fishes, whether the tail is long or relatively short, the pelvis is usually located closer to the head than to the tail tip.
In our description of pelvic shifts, so far we have used the vent as if it is a fixed point of reference on the bodies of all fishes. However, we have already pointed out that even the vent itself has shifted in certain bony fishes. This is shown by the following examples.
In the American knifefishes, the vent and anal fin have shifted forward to a position immediately below the throat. We know of no pelvic girdle or pelvic fins in these fishes and we suspect that they have been squeezed out altogether.
In the priapiumfishes, both the pelvic girdle and the vent have shifted to the throat and the cloaca has been turned inside out, as it were. The faecal, urinary, and genital orifices are laid out separately on a kind of pelvic undercarriage to an otherwise unremarkable minnow-like body.
Finally, the pirate perch, a common freshwater fish in eastern North America, is the most remarkable of all because in this case the vent, normally located just in front of the anal fin, has actually passed ‘through’ the pelvic girdle to an anterior position. The pirate perch is similar to most species of bony fishes in the arrangement of its pelvic fins next to its pectoral fins (our pelvic shift three) and its vent takes the normal position in juveniles. However, by the time of sexual maturity the vent has shifted all the way to a position between the throat and the pelvic fins. The adaptive value of this change seems to be related to depositing its clutch in a safe position. This is because the pirate perch lays its eggs among matted roots underwater and approaches the root-mat head-on rather than turning around – the head being better-designed than the tail to force entry into the tangle. In its peculiarity, the pirate perch perversely restores the original proximity between the vent and the pelvic fins that we saw in sturgeons and cartilaginous fishes – although the excreta now emerge right beneath the gills.
As we have seen, the ventral side of the body of fishes seems to act as a kind of conveyor belt for the pelvic girdle and even the vent – with both being extremely mobile through evolution and taking up positions that suit the particular lineage or the species in question. We close by challenging readers to find a species of bony fish that has its pelvis located closer to its tail tip than to its head (sturgeons certainly come close). In technical terms, bony fish means Osteichthyes, pelvis means the centre of the puboischiac bone or plate, tail tip means the end of the last caudal vertebra (not the tip of the caudal fin), and head means the posterior edge of the gill cover (operculum). Zoological researchers may also find the following hypothesis stimulating – at least as a unifying theme in what can otherwise be a bewilderingly diverse group of animals: “Bony fishes generally differ from cartilaginous fishes in being capable of greater acceleration and deceleration.” Meanwhile, for the less technical among us, a shift of attention across the waistline may help us to see the true breadth of the bodies of fishes, whatever personal significance we attach to our finned fellow creatures.
Should you have any comments or ideas regarding this Biological Exposition, please post a comment in the discussion forum, below.