
In Australia there are many types of plants dispersed and sown by ants. The seeds of these plants have a small but nutritious food-body which is eaten by the ants. The consumption by the ants benefits the plants because the ants usually gather the seeds and bury them in their nests before removing the expendable food-body. This results in the intact seeds being in effect sown – a service particularly valuable to plants adapted to regenerate vigorously after wildfire, because the seeds are buried too deep for them to suffer scorching. Certain lycaenid butterflies are mutualistic with some of the same genera of ants that burythe seeds – their caterpillars being protected by the ants, rewarding them with secretions rich in sugars and amino acids.

Figure 1. Aphaenogaster fulva workers with an eliaosome bearing seed. Such seeds are often buried in ant nests, remaining there intact after the expendable food-body has been consumed. [Photo © Alex Wild]
Here’s where it starts to get intriguing: a comparison of plant lists shows that many of the ant-sown species are also food-plants for lycaenid caterpillars.
From the tangle of relationships arises a new possibility: that certain species of ants may be mutualistic with both their food-plants and the caterpillars of lycaenids which eat these plants. If true, this would be a nice example of resource management by social insects.

Figure 2. Acacia pycnantha, the official floral emblem of Australia, showing the expendable food-body on the seed. Ants use the food-body as a handle to carry the whole item into ant nests, thereafter eating the food-body. Because the seed is hard, smooth, and difficult for the ants to grasp, it remains buried in a viable state, awaiting the next wildfire as a cue to germination. [Photo © Steve Hurst]
Although ants are known to use extrafloral nectaries as well as food-bodies on the seeds of certain types of plants, there are few documented cases of a single species of ant sowing the seeds of its staple food-plant while also ‘farming’ plant-eating insects on the same species of plant, thus deriving food from the same organism in two different ways. Nobody seems to have explored the idea that the same ants may possibly be harvesting both the food-bodies on the seeds and the tasty secretions of lycaenid caterpillars, from a given plant species.
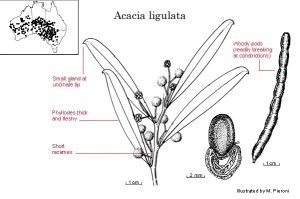
Figure 3. Acacia ligulata, one example of many Australian species in this genus sown by ants which eat the expendable food-body attached to the seed.
There is circumstantial evidence that this complex of reciprocal relationships has evolved independently in parallel on two continents. Fire-prone woody vegetation occurs on nutrient-poor soils in both Australia and southern Africa. Where the vegetation is evergreen but generally unpalatable, the plants most attractive to ant-tended, plant-eating lycaenid caterpillars tend to be nitrogen-fixers regenerating germinatively after fire, nourished by ash. These are also among the plants most likely to be ant-sown by virtue of the food-bodies attached to their seeds.
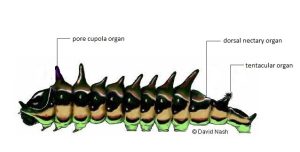
Figure 4. Lycaenid caterpillar typical of species tended by ants, showing the organs used to manage the ants: the pore cupola secretes amino acids, the dorsal nectary organ secretes sugar, and the tentacular organ emits chemical signals to which ants respond. [© David Nash]
Caterpillars of the exclusively Australian lycaenids Jalmenus, Pseudalmenus, Hypochrysops, Nacaduba and Theclinesthes eat the foliage of various species of Acacia, and are tended mainly by various species of ants in the genus Iridomyrmex. These plants are ant-sown nitrogen-fixers, recovering promptly after fire. They are among the more productive and – at least for insects – palatable plants to be found on the nutrient-poor soils of southern Australia.
Now let’s cross the Indian Ocean to another southern continent, to examine what seems almost like a mirror-image of the same complex relationships.

Figure 5. An ant tending the leaf-eating caterpillar of a lycaenid, which secretes foods for its protectors in a mutualism similar to the relationship of many sap-sucking Hemiptera with ants. Photo in the public domain.
Various species in the lycaenid genera Aloeides, Chrysoritis and Tylopaedia have caterpillars that eat Aspalathus, another species-rich genus of legumes but in this case restricted to southern Africa. Like Acacia in Australia, Aspalathus consists mainly of nitrogen-fixing plants which recover conspicuously after intense fires. And the lycaenid Aloeides approximately coincides with the plant Aspalathus in biogeographical terms: it is virtually restricted to Africa south of the equator, and most of its species are restricted to southern Africa. Aloeides is tended by ants of not the genus Iridomyrmex, which is absent from Africa, but instead the genus Lepisiota. And this ant seems analogous with its Australian counterpart, in sowing the seeds of Aspalathus.

Figure 6. Aloeides pierus, exemplifying South African lycaenids which eat the legume Aspalathus during their caterpillar stage. [Photo © Pieter Cramer]
The most familiar example of Aspalathus is rooibos tea, a wild plant that has recently been drafted into domestication because it makes a fragrant brew.
Marring the cosy reciprocities of mutualism is the fact that the relationship between the leaf-eating caterpillars of lycaenids and their food-plants is essentially a destructive one. However, these caterpillars double as predators, frequently eating sap-sucking insects that pester their food-plants. Since Acacia has extrafloral nectaries attractive to ants, and lycaenid butterflies may choose ant-infested plants on which to lay their eggs, one interpretation is that the plant indirectly invites occupation by lycaenid caterpillars despite the price it pays in the greenstuff eaten by these caterpillars.
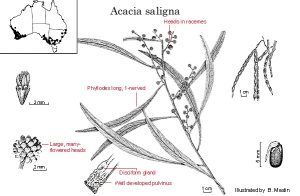
Figure 7. Acacia saligna, another example of an Australian legume sown by ants based on a mutualistic relationship in which the ants consume the expendable food-body on the seed once the seed is buried safely in the ant nest.
Presumably the plant suffers smaller losses to herbivory under the regime of the ant-caterpillar alliance than it would otherwise suffer. And although it’s a stretch to suggest that any caterpillar can do its food-plant any direct favours, the caterpillar metamorphoses into the wings of a potential pollinator. Certain lycaenid butterflies are known to visit Aspalathus for nectar. If they actually contribute to the pollination of their food-plants, then there might be a prospect of a full triangle of mutual benefit – even if the butterflies take away something with the other hand by laying their eggs on the same plants.

Figure 8. Flowers and leaves of rooibos (Aspalathus linearis), indigenous to nutrient-poor sandy soils in the southwestern Cape of South Africa and now harvested commercially for the tea made from its leaves. [Photo © Winfried Bruenken]

Figure 9. Lycaena phlaeas, taking nectar from a flower and potentially pollinating the plant. [Photo © KENPEI]
Regardless of the exact trade-offs involved, the evidence suggests that plants, ants and butterflies have ecological counterparts in intriguing parallels on the two continents. Australian Acacia and southern African Aspalathus resemble each other in nitrogen-fixation, post-fire regeneration, pollination by insects, mutualism with seed-sowing ants, and provision of food for lycaenids. Ants of the genera Iridomyrmex and Lepisiota, occurring respectively in Australia and southern Africa, share similar roles relative to lycaenid caterpillars on legumes, and possibly also relative to the ant-sown plants. Jalmenus and Aloeides are only a few of the lycaenid genera tended as caterpillars by ants on the two continents, but they are the ones most strongly associated with ant-sown legumes.
And the ultimate adaptive benefit of these entanglements?
Well, a possibility is that an ant-based mutualism helps both plant and butterfly to survive intense wildfire. The pupae of species of Jalmenus and Pseudalmenus have been recorded in the safe situations of ant-nests and cracks in the ground. The ants continue to protect the pupae even though at this stage of its life-cycle the lycaenid cannot secrete sugar or amino acids and it lives in debt to the ants. Continual protection by the ants might be a particular favour for those species of lycaenid which grow as caterpillars in the green season, spend the dry season as pupae, and emerge as butterflies when rain falls again, thus sheltering underground during the times of greatest risk of wildfire. By protecting the buried pupae as well as the buried, discarded seeds of legumes, the ants may facilitate the post-fire regeneration of both the plants and the caterpillars that eat the plants.
It may prove difficult to confirm this mutualism for any particular triad of species, because so many species of ants interact with Acacia and Aspalathus and the interactions are not necessarily obligate at a species level. For example, the ants tending the caterpillars of the lycaenid genus Chrysoritis belong to a cosmopolitan genus, Crematogaster, not recorded as participating in the seed-burying mutualism – and therefore unlikely to sow the Aspalathus eaten by the caterpillars. However, if reciprocal benefit can be proven for just one triad – consisting of one species of ant, one species of plant, and one species of lycaenid – the existence of a triangular mutualism would be a nice point to notch in the annals of Biology. This would make a benchmark, defining one extreme in the framework of complex – and seemingly improbable – alliances among disparate organisms.

Figure 10. The lycaenid Chrysoritis thysbe, of which the caterpillars eat the legume Aspalathus in the southwestern Cape of South Africa. [Photo © JMK]
Let’s briefly review the history of scholarship here. It’s been known for two centuries that lycaenid butterflies tend to be mutualistic with ants. Only in the last half-century has the seed-sowing mutualism between plants and ants been fully exposed, particularly in the nutrient-poor parts of two southern continents. There are so many genera and species involved in both mutualisms that biologists have been kept busy – perhaps bewildered – just documenting the technicalities. What would perhaps help to clinch one aspect of this tangle is a particular search image in terms of relationships.
A tantalising question awaiting fieldwork is whether the same species – or colony – of ants sows the food-plant as well as tending the lycaenid caterpillar on a given food-plant – whether Acacia in Australia or Aspalathus in southern Africa. Here’s a chance for some versatile ecologist, undaunted by the technicalities of studying three disparate lineages of organisms, to go to the crux of the matter in confirming for the first time a triangular mutualism in biology at a specific level – consisting of an actual triad of species, one an ant, another a plant, and the third a lycaenid, each simultaneously benefactor and beneficiary. From descriptions so far, the ants seem to be the ones in charge of these diffuse, generic relationships. What beckons here at the Bio-edge is the prospect of identifying at least one triad in which each partner is simultaneously trading favours with both of the other members of a triangular mutualism. And if it can be proven, perhaps it will put the plant, as much as the ant, in the position of authority? … See the hidden half of this blog-post by subscribing here