
If we survey the native faunas of the archipelagos of the world, it is birds rather than mammals that have, in general, succeeded on small patches of land isolated by sea. At first glance, this is unsurprising because the birds – or at least their ancestors – could have flown across the water.
If the pattern is for birds to prevail while mammals fail on islands, New Zealand shows this to a fault. For here we have by far the largest islands lacking any terrestrial[1] mammals in the fauna found at first landfall by human sailors. And nowhere does the old explanation, ‘but mammals couldn’t fly’, fall to earth with more of a thud.
The more we think about the historical accident of birds simply happening to reach New Zealand while mammals did not, the more simplistic it becomes and the more muddled in cause and effect. Worldwide, island faunas are so inconsistent that neither idea, sweepstake dispersal nor the simple distinction between flying and non-flying, have really stood the test of time. In the case of New Zealand nobody is satisfied any more by the old, naive theory – but nothing more buoyant has alighted in the minds of biologists.

Could the reality be that, in New Zealand and elsewhere, the tendency to an avian monopoly on many islands is because birds are superior competitors in these ecological situations – as opposed to simply having the good fortune of access? Could the real biogeographical criterion be the relative competitive abilities of the basic body plans of birds and mammals in the habitat in question, regardless of which class happened to reach the place first?
We won’t quibble for now with the absence of mammalian carnivores in New Zealand. Even this archipelago, almost as vast as California, might have been unable to support any predator larger and less mobile than Haast’s eagle[2]. But we do question the absence of herbivorous, insectivorous, and omnivorous mammals, for which niches would logically seem to be available on the two largest islands. The puzzling reality is that even rodents and insectivorous mammals were absent, their place being taken by flightless birds – called ‘ratites’ – of which moa[3], now extinct, and kiwi[4], still extant, are the most prominent.

Mammals arose deep in dinosaur times, and even the eutherian mammals, which we think of as more modern than marsupials and monotremes, first lived as long ago as 100 million years, in the mid-Cretaceous. At least one mammal[5] is known to have lived 115 million years ago in what is now Victoria in southeastern Australia. This is 35 million years before this part of Gondwana split to produce Zealandia[6], the new continent which went on to produce proto-New Zealand by mountain building after most of Zealandia subsided below sea level[7]. So it seems logical to assume that mammals were present in Zealandia all along, but succumbed to some mysterious change in the nature of the environment as Zealandia became proto-New Zealand.
The inference is that what drove mammals extinct on Zealandia as most of it sank was not the quantity of habitat, but its quality as it experienced geological renewal over a limited area. What matters most is not that these are islands, but the types of habitats[8] these islands offered to terrestrial animals. Which has led us to ponder what geological renewal really means to animals.
So why are moa and kiwi not mammals but birds? Simplistic ‘isolation’ is the wrong answer.
And a nail in the coffin of the old theory was the discovery, in 2006, of an obscure fossil mammal dating to about 17 million years ago. This proved that mammals were present in proto-New Zealand after all.
Likewise, the old view that New Zealand is a raft for relics of the ancient flora and fauna of Gondwana has given way to a realisation that most of the plants, and possibly most of the animals, arrived more recently by sea. New Zealand does, after all, lie in the path of that great conveyor belt of the oceans, the Antarctic Circumpolar Current. With the power of the West Wind Drift, this is the most powerful sea current[9] on Earth and links all the southern continents by surface water flowing continually from west to east at about four kilometres per hour.
If the ancestors of moa arrived only long after the breakup of Gondwana – as is now believed by experts – these flightless ‘ratites’ must have swum or rafted to reach New Zealand, undermining the whole assumption that birds had an advantage of flight[10]. Given that swimming or rafting was the mode of dispersal, why were mammals[11] not also transported this way, as is thought to have happened in the case of Madagascar?
So why are moa and kiwi not mammals but birds? Either way, the old explanation is a dodo. It’s untrue that mammals were absent from New Zealand, and it’s untrue that the ancestors of these flightless birds flew to New Zealand. This raises new possibilities: did marsupials once inhabit proto-New Zealand, together with eucalypts and casuarinas, only to disappear with the conversion of a flat landscape to a mountainous landscape some five million years ago?
Maybe it’s time to start thinking more laterally. So we’ll offer a new theoretical framework. If there are sound, testable reasons why birds took the place of mammals in New Zealand, what are they? Let’s follow a nutritional and biogeochemical rationale.
New Zealand is richer than Australia in phosphorus, calcium and most other nutrients, because of the release of freshly weathered and eroded materials by mountain-building. But it’s actually poorer in critical megacatalysts such as selenium, iodine and cobalt. In fact, New Zealand is one of the most deficient countries on Earth in selenium. Farming sheep on both islands depends on prophyllaxis and treatment of deficiency diseases by supplementing selenium, iodine, cobalt and copper. The problem is clearest in southernmost New Zealand, where a notorious risk of sudden death of human infants has been blamed on iodine and selenium deficiencies[12].
This realisation profoundly changes our evolutionary perspective of these islands. Although micronutrients are needed in seemingly negligible amounts, they’re capable of transforming biogeography because they can be limiting even when all other resources are ample. We regard these elements as potential megacatalysts, at levels from individual physiology to the whole ecosystem.
Selenium, iodine and cobalt operate as catalysts pivotal to life. Glutathione, thyroxine and cobalamin contain selenium, iodine and cobalt, respectively, and activate many enzymes. Thyroid hormones regulate all metabolic processes in the bodies of birds and mammals. Selenium is the key element in protection from chemical damage due to oxidants, poisons and microbial infections. It neutralises the hazardous peroxides made by metabolism, as well as organic and inorganic toxins, and underpins the immune system. The more rapid an animal’s activity, reproduction and growth, the greater its need for antioxidants such as selenium.
Selenium, iodine and cobalt are themselves toxic at concentrations normal for other micronutrients; so they’re difficult to store in the body for future use. At the same time, these megacatalysts are particularly prone to continual losses from the body. Balanced nutrition is therefore paramount – but could proto-New Zealand deliver it?
The poverty of New Zealand in megacatalysts begins with its geology: its main rocks are igneous, with few selenium-rich sediments such as organic shales. The predominant rhyolite is poor in selenium, iodine and cobalt compared with basalt and carbonatite, which are scarce on these islands. New Zealand is also relatively rich in manganese and fluorine, which are metabolic antagonists to our three micronutrients.
What’s more, New Zealand lies so far south that selenium and iodine have been further depleted by past glaciations. Cold climates are likely to boost the metabolic rates of endothermic animals, placing greater demands on this deficient resource. Meanwhile, today’s rainy climates and acidic soils leach and bind selenium, minimising its absorption by plant roots. Sheep succeed in New Zealand only by virtue of continual importation of micronutrients for agriculture, because grasses cannot absorb insoluble forms of selenium excreted by large animals, and recycling is inefficient for this reason. What’s more, selenium and iodine are so soluble and volatile that they’re lost in fluid excreta and even in exhaled air.
Even after tectonic uplift replaced part of the otherwise sinking Zealandia with a new cordillera, rejuvenating the supply of macronutrients, the reproduction and growth of herbivorous, insectivorous, and omnivorous animals might have been limited. Even if all the other nutrients are plentiful, poverty of any of the megacatalysts necessitates a slow pace of life and efficient insulation, locomotion, foraging and reproduction.
So why are moa and kiwi not mammals but birds? Let’s investigate whether birds – including the flightless birds called ratites – could be superior to mammals in their adaptations to poverty of our micronutrients. We will indeed find that the biology of the living relatives of moa and kiwi does show various ostensible adaptations to poverty of megacatalysts.
Let’s look first at birds in general, and then ratites in particular.
• Many birds have higher body temperatures than mammals. However, even these birds may metabolise slowly relative to their great internal surface areas and the distances they move. Whether flying, walking or swimming, birds tend to travel farther than mammals per unit of food energy spent.
• The beaks and long, flexible necks of birds allow extremely selective foraging, and a gizzard is more efficient than teeth for the grinding of nutrient supplements swallowed in the form of earth and bones.
• Birds recycle iodine to the thyroid and selenium to the liver and kidneys more efficiently than mammals do. Osmoregulation and thermoregulation are more efficient in ratites than in mammals, which – owing to their losses of iodine in urine and sweat – depend on binding iodine to blood proteins in a way unnecessary in birds.
• Birds may minimise excretory losses because the powerful avian kidneys and lungs can potentially retrieve the micronutrients more efficiently than in like-sized mammals, while the nasal gland and cloacal bursa of birds may eliminate fluorine while recycling selenium and iodine. The optic pecten of birds potentially nourishes the eye, reducing the concentrations of iodine needed in mammalian eyeballs.
• Feathers are likely to be more efficient than hairs in thermoregulation, sparing megacatalysts directly and indirectly. Fur[13] is likely to demand more selenium relative to energy saved by insulation.
• Birds, whether flying or flightless, have complex air cavities throughout their bodies – in plumage, flesh and bones – making a greater internal surface area than in other animals of similar mass. Interconnecting the surfaces[14] promotes retrieval of selenium and other resources. Aerial flight can thus be achieved with thrift in megacatalysts, despite expenditure of energy; but advantages apply even to flightless birds.
• Birds generally have a greater adult lifespan than like-sized mammals[15], which are the shortest-lived animals of their body size. This partly reflects the efficiency of avian immune and antioxidant systems. By replacing adult individuals as seldom as possible, birds retain resources within the population. Even fast-growing birds tend to delay sexual maturity after reaching full size, whereas mammals attempt to reproduce before they reach full size and die after a brief life of frequent reproduction.
• Birds seal nutrients into eggs, demanding extra calcium but avoiding the urinary losses of megacatalysts that are inevitable with circulation to and from the placenta and mammary glands.
Now let’s look at ratites in particular.
• Being bipedal, ratites are likely to be more mobile than comparable mammals. Flightlessness saves resources even while the bird is resting because pectoral muscles don’t need to be maintained. Bipedal walking is also likely to be economical on the steep slopes and in the dense undergrowth of New Zealand.
• The eggs of the largest ratites were more massive than those of any dinosaur. In the absence of terrestrial predators, moa and kiwi could rely on a few large eggs, minimising the cost of incubation by fasting as the emu[16] does today.
• Large ratites use less energy at rest than the average eutherian mammal of the same body mass at similar temperatures, but also less than expected[17] for birds. For example, the ostrich[18] has a slower metabolism and smaller appetite while resting and walking than coexisting antelopes.
• Ratites have the smallest brains known for animals of their body temperature, because avian brain and body mass scale differently in birds and mammals. Maintaining the brain costs 5–10% of basal energy in most mammals, and is sensitive to deficiency of iodine and selenium. Brains are larger in small birds than in small mammals, but smaller in large birds than in like-sized mammals.
• Unlike herbivorous mammals, male ratites take custody of free-ranging juveniles as well as incubating the eggs. Ratites forage for themselves, even as small chicks, whereas mammals depend on megacatalysts in milk, particularly for brain development.
• Female moa were nearly three times more massive than males in the largest species, a finding with no parallel among mammals. Relegation of parental care to diminutive males might allow female moa to roam freely in search of nutrient supplements such as lichens[19], thus maintaining sufficient body mass to afford the costs of eggs.
Given these advantages of the typical avian, and ratite, body plan and life history strategy, it seems reasonable to suggest that an economical approach would have been competitive in New Zealand with its combined richness of macronutrients and poverty of micronutrients. Hence birds instead of mammals?
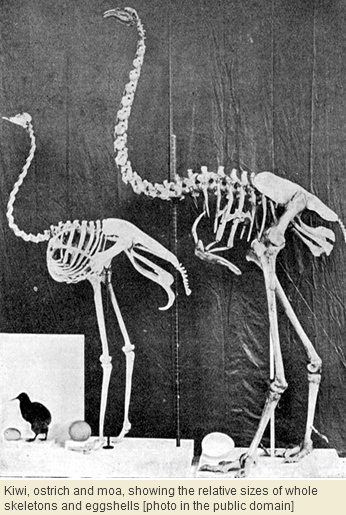
In the light of this new way of thinking, it’s perhaps unsurprising that ratites have also prevailed over mammals in Australia and New Guinea, in a subtle way that has varied over the epochs. At certain times in the past the largest native land animals in New Guinea and Australia have been birds. Furthermore, the largest surviving marsupials – kangaroos – resemble flightless birds in their tendency to bipedality, slow metabolism, small brains, longevity, and embryonic offspring incubated outside the body.
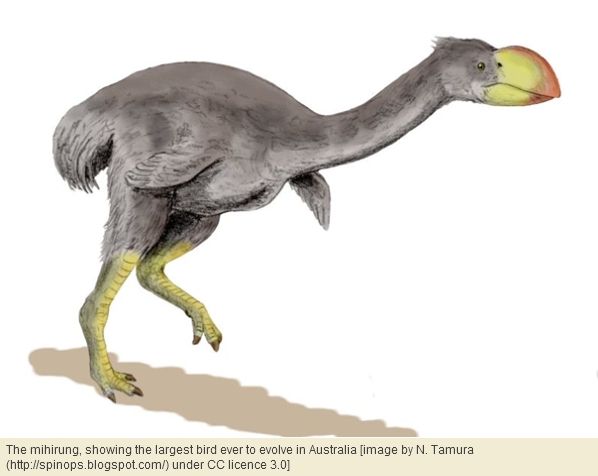
In the Miocene, the largest land animal known from Australasia was a giant bird, the mihirung[20]. Admittedly, marsupials superseded birds by becoming gigantic in the Pleistocene. However, this relationship reverted during the last 30 thousand years: in New Guinea these large marsupials died out while large birds survived, and in Australia kangaroos were reduced to a size inferior to the largest ratites. So, in a subtle way, Australia and New Guinea echoes New Zealand in that ratites have superseded mammals over time.
Notwithstanding the fact that New Guinea has a rich mammalian fauna in contrast to New Zealand, the largest native[21] land animal in New Guinea is the southern cassowary[22], which weighs up to 85 kg and is second only to the ostrich in body mass among living birds. Our point is that ratites successfully competed with mammals in New Guinea, which is similar in size and terrain to New Zealand and also somewhat poor in megacatalysts.

Australia, unlike New Zealand and New Guinea, is so flat and geologically senile that even macronutrients are poor over vast areas. However, even in Australia there is a case to be made for particular deficiency in megacatalysts, in view of the particular effects of pronounced cycles of drought, flood and intense fire on the availability of these micronutrients. For whatever reason, the poverty of large predators in Australia conforms more to an island than to a continent. With this ecological regime, it’s perhaps understandable that ratites have prevailed over mammals to become the largest terrestrial animals among the modern native fauna.
A caveat in discussing Australia: Comparisons of body sizes between birds and mammals must allow for the fact that females are larger than males in Australasian ratites. Although the fully mature male red kangaroo[23] exceeds the largest cassowary[24] so far weighed in Australia, kangaroos are on average much smaller than this because both sexes reach full size only in middle age, and most adult male kangaroos die before reaching full size. Female kangaroos far outnumber males[25], and are below half the body mass of the female southern cassowary at sexual maturity. The average adult female emu[26] is more massive than any female kangaroo alive today and the average adult female southern cassowary[27] in Australia is far more massive than the largest female red kangaroo[28] in full maturity.
With this caveat in mind, we can see that, in a sense, Australia is like New Zealand in its supremacy of birds over mammals. The emu[29] rivals any average for the red kangaroo calculated by weighing the majority of small adults against the few fully mature male kangaroos. So, by our interpretation, the emu is the largest animal native to most[30] of Australia.

So why are moa and kiwi not mammals but birds? We now have the beginnings of a theory based on ecology rather than mere chance.
One of the reasons why biologists have been confused by ratites is their success both on islands, such as New Zealand and Madagascar, and in the heartland of predators: Africa and Asia. However, this apparent paradox can be explained within our conception of ratite ecology, as follows.
Under regimes of frequent predation, such as in Africa and Asia, the inherent efficiency of ratites has been turned from overall thrift to an extreme compensation in reproduction. The ostrich replaces juveniles not just more rapidly than moa did, but at rates unmatched by co-existing like-sized antelopes. In a parallel twist, it also converts its inherent locomotory efficiency to speed by extending the lower limbs. This allows it, like flying birds, to survive regimes of heavy predation by fleeing faster and more enduringly than any antelope. Hence the ostrich has a winning combination: it can grow as rapidly as any like-size mammal despite being long-lived, and it can produce enough eggs to compensate for heavy losses – all despite retaining the typically slow metabolism of ratites. However, all this is possible only by virtue of supplementation[31] at natural mineral licks of the megacatalysts inevitably needed by eggs and growing juveniles – something which is possible for a particularly mobile animal in Africa and Asia.

So why are moa and kiwi not mammals but birds? The adaptive economy of energy and micronutrients in ratites deserves further study. Birds may not be able to spare megacatalysts, overall, when continental predation demands rapid flight and reproduction. But large birds on archipelagos may have superseded marsupials where a measure of relaxation from predation was combined with a regime of subtle nutritional limitation – as on the Zealand that rose, new, from a sinking Zealandia.
—
Acknowledgements: We thank Michael J. Bamford and Martin V. Fey for their generous discussion during the writing of this blog-post.
***
All text and images appearing in this blog are subject to copyright, except those images explicitly stated to be in the public domain. You are not free to use any photographs, for any purpose, without receiving written permission from the copyright holder.
1 except for several species of bats
2 Harpagornis moorei
3 Dinornithiformes
4 Apterygiformes
5 Ausktribosphenos ayktos
6 Zealandia, about half the size of modern Australia, became isolated by broad seas about 65 million years ago, far later than in the case of Madagascar which finally split from India about 88 million years ago. Tectonic renewal of part of Zealandia, to form New Zealand, occurred about 25 million years ago.
7 The land area of Zealandia shrank to as little as 18% of the current size of New Zealand, about 22 million years ago.
8 The fact that recently introduced mammals, such as ruminants and a possum, have thrived in New Zealand does not necessarily prove that niches were formerly empty, because their survival may not ultimately be sustainable without human aid, direct and indirect.
9 This current contains more than 100-fold the volume of all the rivers on Earth and is most powerful south of Tasmania, where it potentially drives flotsam from Australia to New Zealand with particular force.
10 It is now thought that kiwi reached New Zealand even later than this, presumably also by swimming or rafting.
11 Bats show clearly that ancestral ability to fly is a poor explanation for the composition of the New Zealand fauna, which consists of fewer than six species in contrast to the 88 species of bats native to New Guinea.
12 Also possibly relevant to selenium deficiency is the failure of moose (Alces alces), introduced to the southernmost part of the South Island, to thrive in apparently suitable habitat.
13 including wool
14 i.e. where biological reactions and transactions occur
15 except for e.g. bats and primates
16 Dromaius novaehollandiae
17 The basal metabolism of ratites is only 60% of the rate expected for non-passerine birds of their body mass.
18 Struthio camelus
19 Lichens tend to be enriched in iodine owing to absorption directly from air and rain.
20 Dromornis stirtoni
21 excluding the human species
22 Casuarius casuarius
23 Megaleia rufa, which is known to reach a maximum of 85 kilograms in mature males
24 74 kilograms
25 Females outnumber males in the populations of the ratites and marsupials alike.
26 body mass about 38 kilograms
27 i.e. at least 55 kilograms
28 35 kilograms
29 body mass up to 69 kilograms
30 i.e. excluding the small area of fire-free forest in northeastern Queensland, devoid of both emu and large species of kangaroos, in which the southern cassowary occurs
31 The domestic ostrich, like the pig (Sus scrofa), needs twice the average dietary concentration of selenium required by ruminants and equids. This can be explained partly by the extremely rapid reproduction of the ostrich compared to both mammals and island ratites such as moa.